![]()
![]()
Zhaowei ZhuI; Zhenfei FangII; Xinqun HuII; Shenghua ZhouIII
DOI: 10.5935/1678-9741.20150033
BMPR2: Bone morphogenetic protein receptor type II
BMSCs: Bone marrow stromal cells
CTEPH: Chronic thromboembolic pulmonary hypertension
HGF: Hepatocyte growth factor
IGF: Insulin-like growth factor
MSCs: Mesenchymal stem cells
PH: Pulmonary hypertension
RV: Right ventricle
INTRODUCTION
Pulmonary hypertension (PH) is a devastating and refractory disease which is defined by a resting mean pulmonary artery pressure at or above 25 mmHg[1]. Untreated chronic PH can cause a hemodynamic and pathophysiological vicious cycle leading to right ventricle (RV) failure and despite modern treatments, the 3-year survival remains less than 60%[2]. Although currently there is no cure for this disease, treatment has been improved during the past decade, offering both relief from symptoms and prolonged survival. Recently, the regenerative method and gene therapy have been introduced to break the vicious cycle of PH. For example, transplantation of bone marrow-derived mesenchymal stem cells (MSCs) is emerging as a regenerative method to treat PH[3,4]. However, current evidence indicates that the efficacy of MSCs transplantation was unsatisfactory, due to the poor viability and massive death of the engrafted MSCs in the injured tissue. MicroRNAs are short endogenous, conserved, non-coding RNAs and important regulators involved in numerous facets of pathophysiologic processes. There is an obvious involvement of microRNAs in cell differentiation, neovascularization, apoptosis, and others. Nevertheless, the relationship between MSCs, microRNAs and PH is not clear. Here we review the recent findings in MSCs- and microRNAs-based PH treatment, focusing on the potential role of microRNAs regulated MSCs in PH.
MSCS AND PH
MSCs are multipotent progenitor cells that were originally identified in the bone marrow stroma. MSCs have several favorable features for the transplantation therapy of pulmonary hypertension. Besides the ease of isolation and expansion in culture and their capacity to differentiate into multiple lineages, MSCs: have been shown to migrate to sites of injury; they have key interactions with the immune system and generate strong paracrine effects[5]. In addition, Firth et al.[6] identified that a myofibroblast cell phenotype arising from transdifferentiation of differentiation of mesenchymal progenitor cells is predominant within endarterectomized tissues, contributing extensively to the vascular lesion/clot. These properties and findings make MSCs treatment a novel and promising approach for protection from and repair of PH. Recently, A number of animal studies taking use of monocrotaline or hypoxia induced animal model in pulmonary medicine have demonstrated that naive or gene-modified mesenchymal stem cells from bone marrow can ameliorate some of the symptoms of pulmonary hypertension. More interesting, both intratracheal and intravenous administration of MSCs can attenuate pulmonary hypertension in the aspects from endothelial dysfunction[7], alveolar loss and lung inflammation[8] even to ventricle remodeling[7,9-11].
Further researches using gene-modified mesenchymal stem cells treatment also seem successful. Recent studies have found that eNOS[12] or prostacyclin synthase[13] or lung-specific HO-1[14] modified MSCs can not only offer ameliorating effects on PH-related RV impairment but also improve the prognosis and even survival time in PH animals. Although haven't been applied to PH in clinic, all the studies above really provide us a hopeful prospect of MSCs transplantation therapy for PH.
However, the mechanisms of MSCs' therapeutic efficacy are still unclear. Although a robust protection against lung injury on MSCs treatment was observed in most of the above-mentioned animal models, only a small fraction of administered MSCs were detected in the wall of the pulmonary vessels[15]. This observation suggested that engraftment and direct tissue repair were not the sole mechanisms of MSC therapeutic function, and paracrine mechanisms were contemplated.
It is known that MSCs can be mobilized from the total pool of bone marrow stromal cells (BMSCs) when influenced by hypoxia or other injury factors[16]. After mobilization, it can localize into the injured tissue, and even few MSCs can fuse with cells from the host[10]. In addition to being mobilized into the circulation, MSCs have been shown to increase production of growth factors, such as VEGF, insulin-like growth factor (IGF), and hepatocyte growth factor (HGF), when under stress by TNF or hypoxia[17,18]. It is possible that transplanted MSCs may repair injured vascular endothelium by an action involving the release of factors that improve endothelial function or stimulate vascular growth in the injured lung[19,20], which can be partly confirmed by the inhibition of lung inflammation after systemic delivery of MSCs-conditioned media[8]. So, mechanisms for this protection may be not limited to tissue repair, such as engraftment and differentiation of MSCs into specific lung cell types, but also include paracrine factors[7,10,21]. Considering the few numbers of MSCs located in injury tissue, MSCs paracrine signaling maybe a primary mechanism accounting for the beneficial effects of MSCs on responses to injury such as PH.
Among all the paracrine types of MSCs, exosomes, as mediators of cell-cell communication, provide a novel insight into the efficient role of MSCs in PH[11]. Exosomes are a kind of better-defined subclass of secreted membrane microvesicles, which are usually 30 to 100 nm in diameter. They have been isolated and characterized from various cell types, including MSCs. Recent study found MSCs-derived exosomes can exert a pleiotropic protective effect on the lung and inhibit pulmonary hypertension through suppression of hyperproliferative pathways, including STAT3-mediated signaling induced by hypoxia[22].
MICRORNAS AND PH
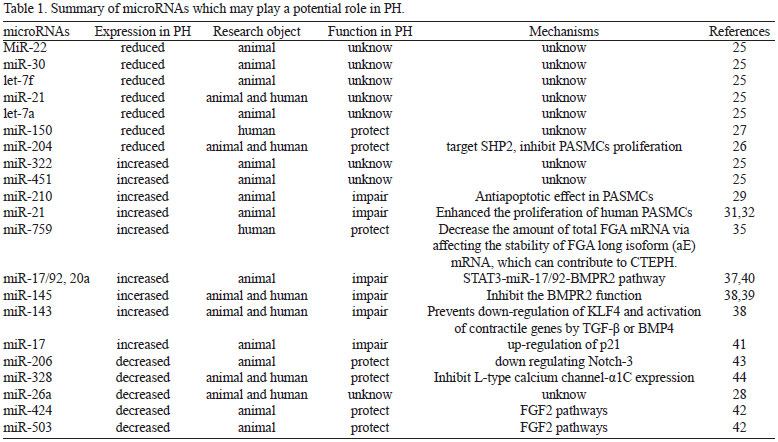
MicroRNAs (miRs) are small, non-coding RNAs regulating gene expression at the post-transcriptional level by mRNA degradation or translational repression[23]. The human genome has been estimated to contain up to 1000 miRNAs[24]. Many miRNAs exhibit a tissue-specific distribution and they appear to play a key role in cell function both under physiological and pathological conditions. Lots of in vivo and in vitro experiments related with functions of microRNAs in PH have emerged recent years. From animal experiments to clinical trials, microRNA expression profiles in PH have been revealed. A range of miRNAs are dysregulated in the lungs of rats exposed to chronic hypoxic and the monocrotaline model of PAH[25]. MiR-22, miR-30 and let-7f were down regulated, whereas miR-322 and miR-451 were up regulated significantly during the development of PH in both hypoxic and monocrotaline models. miR-21 and let-7a were significantly reduced only in monocrotalinetreated rats. miR-204 was consistently down regulated in pulmonary artery smooth muscle cells (PASMCs) from patients with PH and in cells from mice with PH[26]. Besides, circulating miR-150 and miR-26a levels are reduced in patients with poor survival in PH[27,28]. All the evidences above indicate a potential role of microRNAs in PH (Table 1).
As we all know, hypoxia is an important pathogenesis in PH. Inductions of microRNAs can be observed in SMCs cultured with hypoxia and in whole lungs of mice with chronic hypoxia-induced PH. miR-210 is the predominant miRNA induced by hypoxia, which has also been demonstrated by microarray analysis on human in hypoxic PASMCs and in whole lungs of hypoxic mice[29]. The induction of miR-210 is HIF-1α-dependent and triggers anti-apoptotic effects via directly targeting the transcription factor E2F3[29]. Previously study performed on pulmonary artery endothelial cells (PAECs) found that miR-210 can provide an adaptation to hypoxic conditions by targeting Iron-Sulfur Cluster Assembly Proteins 1/2 (ISCU)[30].
MiR-21 can also be induced by hypoxia and overexpression of miR-21 enhanced the proliferation of human PASMCs in vitro and the expression of cell proliferation associated proteins, such as proliferating cell nuclear antigen, cyclin D1, and Bcl-xL, which indicates that miR-21 plays an important role in the pathogenesis of chronic hypoxia-induced pulmonary vascular remodeling[31,32]. Previous study showed that BMP-dependent signaling activation of miR-21 represses Rho-kinase activation in pulmonary artery endothelial cells, thus counteracting the Rho signaling in promoting pulmonary vascular pathology[33]. Besides, miR-21-null mice presented overexpression of RhoB and hyperaction of Rho-kinase activity accompanied by exaggerated manifestation of PH[32].
Chronic thromboembolus is another leading cause of severe PH[34]. Chen et al.[35] investigated the involvement of miR-759 in chronic thromboembolic pulmonary hypertension (CTEPH). CTEPH is characterized by persistent pulmonary embolism that increases pulmonary vascular resistance, resulting in pulmonary hypertension and subsequent right ventricular heart failure. The 3'UTR of FGA was found to interact with miR-759, and a 28-bp deletion polymorphism at this site was found to be more frequent in patients with CTEPH.
Further studies have been investigated to elucidate the concrete mechanisms of microRNAs participating in PH those years. As we all know, bone morphogenetic protein receptor type II (BMPR2), a receptor for the transforming growth factor (TGF-) b family, plays an important role both in endothelial and vascular smooth muscle cells and vascular remodeling of the pulmonary arterial circulation[36]. Several Studies have been designed to identify miRNAs that could inhibit the translation of BMPR-II, and members of the miRNA cluster 17/92 and miR-143, miR-145, miR-20a were identified as the potential regulators[37-40]. All these microRNAs inhibit the BMPR2 function, which were confirmed by experiments in either the patient vascular cells or the PH animal model. miR-145 and miR-143, two highly expressed miRNAs in SMCs, have been shown to play a pivotal role in the modulation of SMC phenotype. In particular, their expression is transcriptionally activated by both TGF-β and BMP4 and promotes a contractile phenotype in SMC by targeting the Kruppel-like factor-4 (KLF4)[38].
Besides the research in pathogenesis and mechanisms, there are also investigations in therapy efficacy of microRNAs for PH in animal models. Pullamsetti et al.[41] demonstrated that inhibition of miR-17 improves heart and lung function in experimental PH by interfering with lung vascular and right ventricular remodeling. The beneficial effects may be related to the up-regulation of p21. And recently, Kim et al.[42] found that reconstitution of miR-424 and miR-503 can ameliorate pulmonary hypertension in experimental models through FGF2 pathways.
Although recent studies found that most PH-related microRNAs usually play a negative role in the pathogenesis process of PH, interestingly, there are still some microRNAs which can play a protective role in PH. miR-204 and miR-206 are two well researched microRNAs, both of which are down regulated in PASMCs from patients with PH or in cells from mice with PH. They all paticipate in the SMCs' proliferation and apoptosis and even differentiation. miR-204 was consistently down regulated in PASMCs from patients with PAH and in cells from mice with PAH[26]. miR-204 show a direct influence on PASMC function and delivery of miR-204 mimics to the lungs of mice with PAH significantly can reduce disease severity. miR-206 can alleviate PAH through down regulating Notch-3 expression, which is key a factor in PAH development[43].
Besides, hypoxia produced a significant inhibition of miR-328 expression, which has been identified as a strong candidate responsible for hypoxic pulmonary vasoconstriction. Overexpressing miR-328 in the transgenic mice remarkably decreased the right ventricular systolic pressure and PA wall thickness under both normoxia and hypoxia. Through inhibiting L-type calcium channel-α1C expression the insulin growth factor 1 receptor, ultimately leading to apoptosis of pulmonary arterial smooth muscle cells[44].
POSSIBLE AND NOVEL LINK BETWEEN MSCS AND MIRNAS IN PH
It is well known that miRNAs have been implicated in many processes of stem cell functions, including cell proliferation, differentiation and apoptosis. Recent studies[45] suggest that mesenchymal stem cells have discrete miRNA expression profiles that can account for the intrinsic stem cell properties of self-renewal and pluripotency. Through certain modified microRNAs, up or down regulation, there must be ways to enhance the viability of engrafted MSCs in the injured pulmonary tissue.
Exosomes have emerged as a novel media between kinds of cells. And exosomes based therapy has been confirmed by many researches. In consideration of its microRNAs-carried function, it is feasible to treat with PH by the microRNAs-carried exosomes secreted by MSCs. Recently, this hypothesis has been confirmed in a research that demonstrate MSCs can regulate neurite outgrowth by transfer of miR-133b to neural cells via exosomes[46].
In a word, either mimicking or antagonizing microRNA actions, MSCs functions can be regulated by microRNAs to enhance the properties of cell differentiation or anti-apoptosis. Considering that microRNAs can be delivered by exosomes secreted by MSCs, it is likely that overexpression of special microRNAs like miR-204/206/328 in MSCs will hopefully enhance MSCs therapeutic efficacy for PH. So, microRNAs may be used as novel regulators in MSC-based therapy in PH and microRNAs-regulated MSCs transplantation may represent promising therapeutic strategy for PH patients in the future.
REFERENCES
1. Galiè N, Hoeper MM, Humbert M, Vachiery JL, Barbera JA, Beghetti M, et al. ; ESC Committee for Practice Guidelines (CPG). Guidelines for the diagnosis and treatment of pulmonary hypertension: the Task Force for the Diagnosis and Treatment of Pulmonary Hypertension of the European Society of Cardiology (ESC) and the European Respiratory Society (ERS), endorsed by the International Society of Heart and Lung Transplantation (ISHLT). Eur Heart J 2009;30(20):2493-537. [MedLine]
2. Humbert M, Sitbon O, Chaouat A, Bertocchi M, Habib G, Gressin V, et al. Survival in patients with idiopathic, familial, and anorexigen-associated pulmonary arterial hypertension in the modern management era. Circulation. 2010;122(2):156-63. [MedLine]
3. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature. 2002;418(6893):41-9. [MedLine]
4. Zhao YD, Courtman DW, Deng Y, Kugathasan L, Zhang Q, Stewart DJ. Rescue of monocrotaline-induced pulmonary arterial hypertension using bone marrow-derived endothelial-like progenitor cells: efficacy of combined cell and eNOS gene therapy in established disease. Circ Res. 2005;96(4):442-50. [MedLine]
5. Myers TJ, Granero-Molto F, Longobardi L, Li T, Yan Y, Spagnoli A. Mesenchymal stem cells at the intersection of cell and gene therapy. Expert Opin Biol Ther. 2010;10(12):1663-79. [MedLine]
6. Firth AL, Yao W, Ogawa A, Madani MM, Lin GY, Yuan JX. Multipotent mesenchymal progenitor cells are present in endarterectomized tissues from patients with chronic thromboembolic pulmonary hypertension. Am J Physiol Cell Physiol. 2010;298(5):C1217-25. [MedLine]
7. Baber SR, Deng W, Master RG, Bunnell BA, Taylor BK, Murthy SN, et al. Intratracheal mesenchymal stem cell administration attenuates monocrotaline-induced pulmonary hypertension and endothelial dysfunction. Am J Physiol Heart Circ Physiol. 2007;292(2):H1120-8. [MedLine]
8. Aslam M, Baveja R, Liang OD, Fernandez-Gonzalez A, Lee C, Mitsialis SA, et al. Bone marrow stromal cells attenuate lung injury in a murine model of neonatal chronic lung disease. Am J Respir Crit Care Med. 2009;180(11):1122-30. [MedLine]
9. Guarita-Souza LC, Carvalho KAT, Rebelatto C, Senegaglia AC, Hansen P, Furuta M, et al. A comparação entre o transplante de células tronco mononucleares e mesenquimais no infarto do miocárdio. Rev Bras Cir Cardiovasc. 2005;20(3):270-8. View article
10. Spees JL, Whitney MJ, Sullivan DE, Lasky JA, Laboy M, Ylostalo J, et al. Bone marrow progenitor cells contribute to repair and remodeling of the lung and heart in a rat model of progressive pulmonary hypertension. FASEB J. 2008;22(4):1226-36. [MedLine]
11. Umar S, de Visser YP, Steendijk P, Schutte CI, Laghmani el H, Wagenaar GT, et al. Allogenic stem cell therapy improves right ventricular function by improving lung pathology in rats with pulmonary hypertension. Am J Physiol Heart Circ Physiol. 2009;297(5):H1606-16. [MedLine]
12. Kanki-Horimoto S, Horimoto H, Mieno S, Kishida K, Watanabe F, Furuya E, et al. Implantation of mesenchymal stem cells overexpressing endothelial nitric oxide synthase improves right ventricular impairments caused by pulmonary hypertension. Circulation. 2006;114(1 Suppl):I181-5. [MedLine]
13. Takemiya K, Kai H, Yasukawa H, Tahara N, Kato S, Imaizumi T. Mesenchymal stem cell-based prostacyclin synthase gene therapy for pulmonary hypertension rats. Basic Res Cardiol. 2010;105(3):409-17. [MedLine]
14. Liang OD, Mitsialis SA, Chang MS, Vergadi E, Lee C, Aslam M, et al. Mesenchymal stromal cells expressing heme oxygenase-1 reverse pulmonary hypertension. Stem Cells. 2011; 29(1):99-107. [MedLine]
15. Anversa P, Perrella MA, Kourembanas S, Choi AM, Loscalzo J. Regenerative pulmonary medicine: potential and promise, pitfalls and challenges. Eur J Clin Invest. 2012;42(8):900-13. [MedLine]
16. Rochefort GY, Delorme B, Lopez A, Hérault O, Bonnet P, Charbord P, et al. Multipotential mesenchymal stem cells are mobilized into peripheral blood by hypoxia. Stem Cells. 2006;24(10):2202-8. [MedLine]
17. Wang M, Crisostomo P, Herring C, Meldrum KK, Meldrum DR. Human progenitor cells from bone marrow or adipose tissue produce VEGF, HGF and IGF-1 in response to TNF by a p38 MAPK-dependent mechanism. Am J Physiol Regul Integr Comp Physiol. 2006;291(4):R880-4. [MedLine]
18. Rehman J, Traktuev D, Li J, Merfeld-Clauss S, Temm-Grove CJ, Bovenkerk JE, et al. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation. 2004;109(10):1292-8. [MedLine]
19. Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S, et al. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ Res. 2004;94(5):678-85. [MedLine]
20. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997;276(5309):71-4. [MedLine]
21. Patel KM, Crisostomo P, Lahm T, Markel T, Herring C, Wang M, et al. Mesenchymal stem cells attenuate hypoxic pulmonary vasoconstriction by a paracrine mechanism. J Surg Res. 2007;143(2):281-5. [MedLine]
22. Lee C, Mitsialis SA, Aslam M, Vitali SH, Vergadi E, Konstantinou G, et al. Exosomes mediate the cytoprotective action of mesenchymal stromal cells on hypoxia-induced pulmonary hypertension. Circulation. 2012;126(22):2601-11. [MedLine]
23. Small EM, Olson EN. Pervasive roles of microRNAs in cardiovascular biology. Nature. 2011;469(7330):336-42. [MedLine]
24. Zhang L, Huang J, Yang N, Greshock J, Megraw MS, Giannakakis A, et al. microRNAs exhibit high frequency genomic alterations in human cancer. Proc Natl Acad Sci U S A. 2006;103(24):9136-41.
25. Caruso P, MacLean MR, Khanin R, McClure J, Soon E, Southgate M, et al. Dynamic changes in lung microRNA profiles during the development of pulmonary hypertension due to chronic hypoxia and monocrotaline. Arterioscler Thromb Vasc Biol. 2010;30(4):716-23. [MedLine]
26. Courboulin A, Paulin R, Giguère NJ, Saksouk N, Perreault T, Meloche J, et al. Role for miR-204 in human pulmonary arterial hypertension. J Exp Med. 2011;208(3):535-48. [MedLine]
27. Rhodes CJ, Wharton J, Boon RA, Roexe T, Tsang H, Wojciak-Stothard B, et al. Reduced miR-150 is associated with poor survival in pulmonary arterial hypertension. Am J Respir Crit Care Med. 2013;187(3):294-302. [MedLine]
28. Schlosser K, White RJ, Stewart DJ. miR-26a linked to pulmonary hypertension by global assessment of circulating extracellular MicroRNAs. Am J Respir Crit Care Med. 2013;188(12):1472-5. [MedLine]
29. Gou D, Ramchandran R, Peng X, Yao L, Kang K, Sarkar J, et al. miR-210 has an antiapoptotic effect in pulmonary artery smooth muscle cells during hypoxia. Am J Physiol Lung Cell Mol Physiol. 2012;303(8):L682-91. [MedLine]
30. Chan SY, Zhang YY, Hemann C, Mahoney CE, Zweier JL, Loscalzo J. MicroRNA-210 controls mitochondrial metabolism during hypoxia by repressing the iron-sulfur cluster assembly proteins ISCU1/2. Cell Metab. 2009;10(4):273-84. [MedLine]
31. Yang S, Banerjee S, Freitas Ad, Cui H, Xie N, Abraham E, et al. miR-21 regulates chronic hypoxia-induced pulmonary vascular remodeling. Am J Physiol Lung Cell Mol Physiol. 2012;302(6):L521-9. [MedLine]
32. Parikh VN, Jin RC, Rabello S, Gulbahce N, White K, Hale A, et al. MicroRNA-21 integrates pathogenic signaling to control pulmonary hypertension: results of a network bioinformatics approach. Circulation. 2012;125(12):1520-32. [MedLine]
33. Connolly MJ, Aaronson PI. Key role of the RhoA/Rho kinase system in pulmonary hypertension. Pulm Pharmacol Ther. 2011;24(1):1-14. [MedLine]
34. Jatene FB, Pêgo-Fernandes PM, Poveda S, Monteiro R, Cukier A, Mady C, et al. Tratamento cirúrgico da embolia pulmonar crônica: análise da experiência inicial. Rev Bras Cir Cardiovasc. 1995;10(2):70-6.
35. Chen Z, Nakajima T, Tanabe N, Hinohara K, Sakao S, Kasahara Y, et al. Susceptibility to chronic thromboembolic pulmonary hypertension may be conferred by miR-759 via its targeted interaction with polymorphic fibrinogen alpha gene. Hum Genet. 2010;128(4):443-52. [MedLine]
36. Rabinovitch MJ. Molecular pathogenesis of pulmonary arterial hypertension. Clin Invest. 2012;122(12):4306-13.
37. Brock M, Trenkmann M, Gay RE, Michel BA, Gay S, Fischler M, et al. Interleukin-6 modulates the expression of the bone morphogenic protein receptor type II through a novel STAT3-microRNA cluster 17/92 pathway. Circ Res. 2009;104(10):1184-91. [MedLine]
38. Davis-Dusenbery BN, Chan MC, Reno KE, Weisman AS, Layne MD, Lagna G, et al. down-regulation of Kruppel-like factor-4 (KLF4) by microRNA-143/145 is critical for modulation of vascular smooth muscle cell phenotype by transforming growth factor-beta and bone morphogenetic protein 4. J Biol Chem. 2011;286(32):28097-110. [MedLine]
39. Caruso P, Dempsie Y, Stevens HC, McDonald RA, Long L, Lu R, et al. A role for miR-145 in pulmonary arterial hypertension: evidence from mouse models and patient samples. Circ Res. 2012;111(3):290-300. [MedLine]
40. Brock M, Samillan VJ, Trenkmann M, Schwarzwald C, Ulrich S, Gay RE, et al. AntagomiR directed against miR-20a restores functional BMPR2 signalling and prevents vascular remodelling in hypoxia-induced pulmonary hypertension. Eur Heart J. 2014;35(45):3203-11. [MedLine]
41. Pullamsetti SS, Doebele C, Fischer A, Savai R, Kojonazarov B, Dahal BK, et al. Inhibition of microRNA-17 improves lung and heart function in experimental pulmonary hypertension. Am J Respir Crit Care Med. 2012;185(4):409-19. [MedLine]
42. Kim J, Kang Y, Kojima Y, Lighthouse JK, Hu X, Aldred MA, et al. An endothelial apelin-FGF link mediated by miR-424 and miR-503 is disrupted in pulmonary arterial hypertension. Nat Med. 2013;19(1):74-82. [MedLine]
43. Jalali S, Ramanathan GK, Parthasarathy PT, Aljubran S, Galam L, Yunus A, et al. Mir-206 regulates pulmonary artery smooth muscle cell proliferation and differentiation. PLoS One. 2012;7(10):e46808. [MedLine]
44. Guo L, Qiu Z, Wei L, Yu X, Gao X, Jiang S, et al. The microRNA-328 regulates hypoxic pulmonary hypertension by targeting at insulin growth factor 1 receptor and L-type calcium channel-α1C. Hypertension. 2012;59(5):1006-13. [MedLine]
45. Guo L, Zhao RC, Wu Y. The role of microRNAs in self-renewal and differentiation of mesenchymal stem cells. Exp Hematol. 2011;39(6):608-16. [MedLine]
46. Xin H, Li Y, Buller B, Katakowski M, Zhang Y, Wang X, et al. Exosome-mediated transfer of miR-133b from multipotent mesenchymal stromal cells to neural cells contributes to neurite outgrowth. Stem Cells. 2012;30(7):1556-64. [MedLine]
No financial support.
Authors' roles & responsibilities
ZZ: Study conception and design
ZF: Study conception and design
XH: Analysis and/or interpretation of the data
SZ: Final approval of the manuscript
Article receive on Monday, December 22, 2014
 All scientific articles published at rbccv.org.br are licensed under a Creative Commons license
All scientific articles published at rbccv.org.br are licensed under a Creative Commons license













All rights reserved 2017 / © 2024 Brazilian Society of Cardiovascular Surgery
DEVELOPMENT BY ![]()
 Read in English
Read in English
 English PDF
English PDF
 Print
Print
 Send this article by email
Send this article by email
 How to cite this article
How to cite this article
 Submit a comment
Submit a comment
 Mendeley
Mendeley
 Pocket
Pocket